Acai Beere
Anwendung, Wirkung, Studien | Wo kaufen?
Die Acai Beere (sprich ass-a-i) ist die Frucht der Kohlpalme Euterpe oleracea. Diese Palme wächst in Südamerika auf feuchten, humusreichen Böden besonders im Amazonasgebiet. Bei den indigenen Völkern gehört sie zu den festen Nahrungsmitteln. Diese haben ein großes Wissen um die Frucht und nutzen das Öl der Frucht gegen Diarrhö, einen Aufguss gegen Hautgeschwüre und es findet auch Anwendung bei Fieber.

[amazon table=“2468″]
In Nordamerika und Europa erlangte sie in den letzten Jahren Berühmtheit als angebliches Schlankheitsmittel, doch es steckt etwas ganz anderes in ihr. Durch einen hohen Anteil an Antioxidantien, die sehr gut in den Körper aufgenommen werden, hat die Acai Beere einen großen gesundheitlichen Benefiz für den Menschen. Antioxidantien haben einen positiven Effekt auf den Cholesterinwert, Diabetes, Arthritis, auf Erkrankungen des Herzkreislaufsystems, Arteriosklerose, Bluthochdruck, Fibromyalgie und Immunstörungen. Wissenschaftler der Emory University School of Medicine fanden heraus, dass Fruchtfliegen, die sich von Acai Beeren ernähren, 3 Mal länger leben. Dies ist auf Antioxidantien zurückzuführen, die die Zellen schützen und deren Alterungsprozess verlangsamen.
Der Mix an verschiedenen Inhaltsstoffen führte in den letzten Jahren dazu, dass unterschiedliche Studien zur Auswirkung der Acai Beere auf den menschlichen Organismus durchgeführt worden sind, um das Potential der Beere vollständig zu entdecken (dazu mehr weiter unten im Artikel). Eine Wirkung als Schlankheitswunder konnte in diesen Studien nicht nachgewiesen werden, doch was in der Beere steckt, ist viel mehr wert.
Anwendung & Wirkung
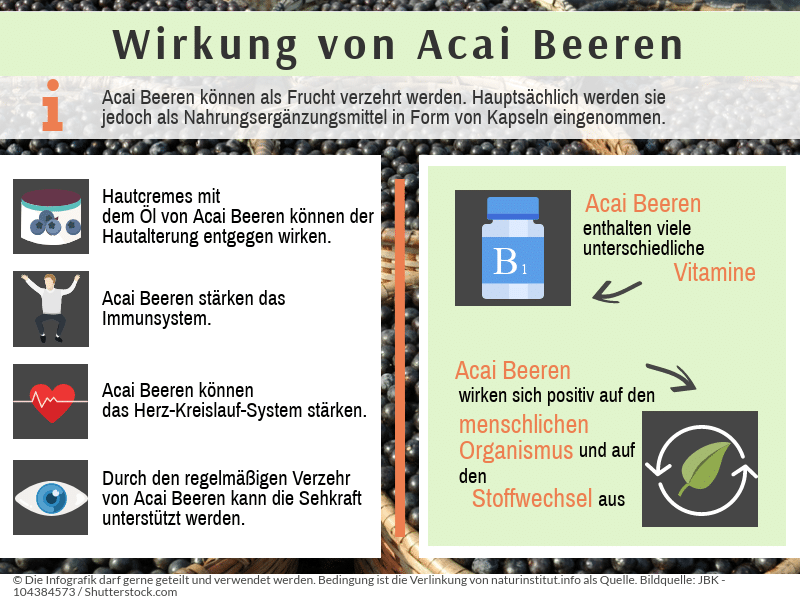
Die Acai Beere ist zum einen Bestandteil in der Kosmetik, speziell in Hautcremes. Die antioxidative Wirkung ist um das 1,5 fache größer als beim Granatapfel. Durch die UV-Strahlung entstehen freie Radikale und werden für die Hautalterung verantwortlich gemacht. Das Öl aus der Acai Beere ist reich an Antioxidantien und Polyphenolen. Es soll der Hautalterung entgegenwirken und spendet zudem Feuchtigkeit. Cremes werden zweimal täglich aufgetragen.
Die Acai Beere kann als Frucht verzehrt werden. Hauptsächlich wird sie als Nahrungsergänzungsmittel eingenommen. Als bekannteste Einnahmeform gelten Kapseln. Die Kapseln besitzen zwischen 1000 und 2000 mg Acai Beeren-Extrakt. Es sollen 5000-6000mg täglich aufgenommen werden. Die Kapseln werden mit Flüssigkeit eingenommen. Eine weitere Form der Einnahme ist Püree aus den Früchten. Es wird ein täglicher Verzehr von 30ml des Pürees empfohlen. Als Saft werden Flaschen zu 500 ml verkauft. Nach dem Öffnen sollte der Saft kühl gelagert und innerhalb von 14 Tagen verbraucht werden. Es gibt die Acai Beere in Pulverform. Die Empfehlung liegt bei ca. 5g des Pulvers täglich.
Neben dem Verzehr von Acai Beeren und der Einnahme als Saft sowie Püree findet sich das Extrakt der Acai Beeren hauptsächlich in Nahrungsergänzungsprodukte wie Pulver oder Kapseln. Das Augenmerk liegt dabei noch auf die Anwendung im Rahmen einer Diät.
In der Kosmetik finden die Extrakte der Acai Beere immer mehr Beachtung durch den hohen Anteil an Antioxidantien. Das Extrakt wird Hautcremes beigefügt, um der Hautalterung vorzubeugen ähnlich wie beim Arganöl .
Acai Beere abnehmen
Acai-Beeren sind tatsächlich echte Superfoods. Sie bietet in konzentrierter Form eine enorme Menge an Vitalstoffen, welche der Gesundheit dienlich sind. Die Bandbreite reicht von Anti-Oxidantien über zahlreiche Vitamine und Ballaststoffe bis hin zu Stoffen, welche die Venen und Arterien von Ablagerungen befreien können. Dazu können übrigens alle essbaren, dunklen Beeren gezählt werden: Es ist der dunkle, natürliche Farbstoff, welcher die Cholesterin-Plaques in den Blutgefäßen abtragen kann. Darum wird auch der Rotwein immer so sehr von den Herz- und Gefäßspezialisten empfohlen: Der Alkohol weitet die Gefäße und der rote Farbstoff trägt die Ablagerungen ab.
Eine dezidierte Wirksamkeit von Acai-Beeren in Bezug auf das Abnehmen besteht leider nicht. Allerdings sollte man sich als Abnehmwilliger grundsätzlich von der Idee verabschieden, dass dazu nur irgendein Wundermittel notwendig sei. Abnehmen ist im Grunde ein ganz simpler mathematischer Prozess: Werden am Tag mehr Kalorien aufgenommen als verbraucht, nimmt man zu. Nimmt man weniger Kalorien auf als man verbraucht, nimmt man ab – das ist eigentlich die ganze Kunst.
Für ein erfolgreiches Abnehmen sind drei Dinge nötig:
- Allgemeines Wissen
- Kenntnis des eigenen Körpers
- Disziplin
Das allgemeine Wissen ist erforderlich, um die Zusammenhänge zwischen Kalorienaufnahme und Aufbau von Fettpolstern zu verstehen. Dickmacher sind alle Nährstoffe, die zu Energie verstoffwechselt werden können. Je einfacher die Energie verfügbar gemacht wird, desto eher handelt es sich um ein dick machendes Lebensmittel.
Die Energielieferanten sind, geordnet nach ihrer einfachen Verstoffwechselbarkeit:
- Kohlenhydrate: Weißes Mehl, gekochte Kartoffeln, Zucker, Backwaren, Nudeln
- Fette: Öle, Schmalz, Butter
- Eiweiß: Fleisch, Fisch, bestimmte eiweißhaltige Gemüsesorten
Die Natur hat es so eingerichtet, dass energiereiche Nährstoffe besonders gut schmecken. Der Mensch hat dabei ein Produkt entwickelt, welches es in der Natur praktisch nicht gibt: Die Kombination aus fetthaltigen und Kohlehydrat reichen Produkten. Diese Kombi-Lebensmittel sind es, welche nicht nur besonders gut schmecken – sie machen auch ganz besonders schnell dick. Leider sind es gerade diese verarbeiteten Produkte, die sich besonders gut verkaufen. Zu ihnen zählen:
- Backwaren
- Nudeln mit Soße
- Kekse, Schokolade
- Eiscreme
- Schokoriegel
Die Kombination aus „Süß“ und „Fettig“ ist dem Gehirn praktisch unbekannt. Es spricht daher unseligerweise das Suchtzentrum stark an, da es schnell viele und wohlschmeckende Kalorien bietet.
Das Gehirn ist leider immer noch darauf programmiert, einen Hungertod um jeden Preis abwenden zu müssen. Das führt zu dem irrationalen Essverhalten, das wir heute haben.
Es ist daher wichtig, sich stets ganz genau darüber bewusst zu sein, was man gerade ist. Idealerweise wird ein „Kalorien-Tagebuch“ geführt, in dem die Summe aller aufgenommenen Dickmacher genau erfasst wird.
Kenntnis des eigenen Körpers
Theoretisch kann man so viele Kalorien aufnehmen, wie man will – wenn die ganze Energie sofort wieder abtrainiert wird, nimmt man nicht zu. Doch hier kann man beruhigt sein: Es muss nicht jeder einzelne Bissen abtrainiert werden. Einen Großteil der täglich zugeführten Energie verbraucht der Körper über den so genannten „Grundumsatz“. Dies ist die Energiemenge, die ein Körper pro Tag zum Erhalt seiner Funktionen selbst verbraucht. Der Energieverbraucher Nr. 1 ist das Gehirn, gefolgt vom Herz-Kreislauf-System und allen anderen Stoffwechselprozessen.
Der Grundumsatz ist sehr individuell. Im Internet gibt es Rechner, mit denen man sich seinen Grundumsatz berechnen lassen kann. Ein übergewichtiger Mann mittleren Alters und geringer sportlicher Betätigung hat bereits einen Grundumsatz von 2600 Kalorien. Da aber nicht jede Kalorie auch wirklich verstoffwechselt sondern viele wieder ausgeschieden werden, muss dieser Mann 3600 Kalorien pro Tag aufnehmen, um den Grundumsatz abdecken zu können.
3600 Kalorien sind schon eine ganze Menge. Allerdings: Ein Schokoriegel alleine hat schon 500 Kalorien. Meistens sind es aber gerade diese Zwischendurch-Snacks, die den größten Teil eines Übergewichtes ausmachen.
Wie gesagt, Acai-Beere ist jetzt nicht unbedingt dafür bekannt, dass man damit besonders schnell und nachhaltig abnehmen könnte. Jedoch kann die Auseinandersetzung mit dieser Superfood-Beere sehr gut dazu beitragen, sich seines Ernährungsverhaltens bewusst zu werden. Sie kann damit eine Eintrittskarte für ein neues, besseres Körpergefühl sein. Dazu gehört allerdings auch eine regelmäßige und ausgewogene Bewegung. Das muss gar nicht mal so viel sein: 30 Minuten Spazieren gehen am Tag reichen schon völlig aus, um eine Verbesserung des Stoffwechsels anzuregen. Dann überlegt man sich den Schokoriegel danach aber zwei Mal: Will man sich wirklich die gerade abgelaufenen Kalorien wieder draufpacken – oder bekämpft man den Zuckerhunger doch besser mit einer Handvoll Acai-Beere?
Ohne Disziplin geht es nicht
Der Wille zum Abnehmen ist das größte Hindernis auf dem Weg zum Ziel eines schönen, schlanken Körpers. Hier spielen dem Übergewicht gleich mehrere Faktoren in die Hände, die es dem Betroffenen stets so schwer machen.
Zunächst ist da die Gewohnheit. Um sich eine neue Lebensweise anzugewöhnen braucht es ca. 6 – 10 Wochen. In dieser Zeit muss man eisern um jeden Bissen und jedes bisschen Bewegung kämpfen. Doch dann ist es meistens geschafft. Der Körper und der Geist haben die neuen Muster angenommen und ab dann geht es mit dem Abnehmen immer leichter.
Ein Riesenproblem beim Abnehmen sind die Endorphine. Zwar gibt es tatsächlich das „Runner´s High“ – den minuten- bis stundenlangen berauschenden Glückszustand bei starker körperlicher Anstrengung. Jedoch erreicht man diesen Zustand erst nach vielen, vielen Stunden des harten Trainings und mit der Eliminierung fast jeder Fettzelle im Körper. Fett hat in Bezug auf das Endorphin leider eine verheerende Wirkung: Es wirkt wie ein Filter, welches die Endorphin-Hormone im Blut abfängt und nur vermindert zum Hippocampus durchlässt. Das bedeutet: Übergewichtige müssen sich doppelt anstrengen und werden noch nicht einmal dafür belohnt. Man kommt schwitzend und keuchend vom Sport – fühlt sich aber keineswegs stolz oder glücklich dafür. Um dann doch noch an seine Endorphine zu kommen, „Belohnen“ sich viele Abnehmwillige später mit einem Stück Kuchen, einer Pommes-Frites oder einer Tafel Schokolade. Die wird dann wieder mit Frustration gefolgt – und der innere Schweinehund ist für die nächste Sportrunde wieder ein Stück stärker geworden.
Hier kann man nur empfehlen durchzuhalten. Je mehr Fett abgebaut wird, desto besser kommen die Endorphine dort an, wo sie sollen. Nach sechs Wochen ist die Gewöhnung erreicht. Ab dann geht es erfahrungsgemäß schneller mit dem Abnehmen. Bis dahin sollte man sich ausschließlich gesunde Snacks gönnen – zum Beispiel Acai Beere.
Studien
Durch die Anwendung der Acai Beere in der Nahrungsmittelindustrie hat das Interesse an der Beere als Studienobjekt in den vergangenen Jahren stark zugenommen und zu den einzelnen Inhaltsstoffen können inzwischen Nachweise über ihre Effekte geliefert werden.
Einige Forschungsarbeiten wurden an der Universität von Florida, Institute of Food and Agriculture Sciences unter Stephen Talcott und seinem Team durchgeführt. 2006 konnte er an Zellkulturen nachweisen, dass das Extrakt der Acai Beere eine Selbstzerstörungsreaktion in bis zu 86 % der vorliegenden Leukämiezellen auslöste. Es wird jedoch ausdrücklich darauf hingewiesen, dass dies nicht bedeutet, dass eine Leukämie verhindert werden kann. Es gibt keine Zahlen dazu, wie es sich bei Leukämieerkrankungen verhält. Wenn das Extrakt in den Körper gelangt, sollte bedacht werden, dass Verdauung, Absorption und chemische Prozesse mit eine Rolle spielen.
Daher untersuchte Stephen Talcott 2008 mit seinem Team, wie die Antioxidantien aus der Acai Beere in den menschlichen Organismus aufgenommen werden. Es konnte gezeigt werden, dass beim Verzehr der Acai Beere die antioxidative Aktivität im Blut anstieg.
2011 fand eine Forschergruppe der Universität Kobe und Fukui, Japan, heraus, dass die Acai Beere eine hemmende Wirkung auf die IgE-vermittelte Aktivierung der Mastzellen hat. Diese Aktivierung spielt eine große Rolle bei allergischen Reaktionen.
Alle Forschungsgruppen sind sich einig, dass weitere Studien wichtig sind, um das gesamte Potential der Acai Beere erklären zu können.
Inhaltsstoffe
Bei der Einnahme von 100 g Acai Beeren nimmt der Körper 80 bis 250 Kalorien zu sich, je nach Art der Zubereitung. 50 % der gesamten Energie kommt von Fetten. Der geringe Zuckeranteil liegt bei 3,6 %. Der Geschmack wird als fettig-erdig bezeichnet, zum Teil auch adstringend (die Haut zusammenziehend). Daher wird ein Produkt der Acai Beeren mit Saft oder Sirup gemischt vor der Einnahme.
Der wichtigste Inhaltsstoff der Acai Beere sind die Anthocyane Cyanid-3-glucosid und Cyanid-3-glucosid-Cumarat: Anthocyane sind Antioxidantien. Sie fangen freie Radikale im Organismus ab und inaktivieren diese reaktiven Verbindungen. Bei den freien Radikalen handelt es sich um hochreaktive Sauerstoffverbindungen. Sie werden im Körper gebildet. Schadstoffe in der Luft, Chemikalien und auch UV-Strahlung kann die Entstehung noch verstärken. Sie verursachen im Organismus einen oxidativen Stress, der zu Zellschäden führen kann. Diese werden in Verbindung mit der Zellalterung und damit auch mit der Entstehung verschiedener Krankheiten gebracht. Durch das Abfangen der freien Radikale im Organismus wirken die Anthocyane entzündungshemmend, antiproliferativ, zytoprotektiv (zellschützend) und immunstimulierend, d.h. das Immunsystem sowie das Herz-Kreislauf-System werden gestärkt, die Sehkraft wird unterstützt und der Alterungsprozess wird verlangsamt.
- Auf 100g kommen 133-309 mg Calcium: Calcium ist der am stärksten vorkommende Mineralstoff im menschlichen Körper und findet sich hauptsächlich in den Knochen und Zähnen. Gespeichert ist es im Knochen. Besteht ein Calciummangel, wird es aus dem Knochen gelöst, welcher dadurch an Stabilität verlieren kann. Dies kann im Alter zum Krankheitsbild der Osteoporose führen. Doch neben der Stabilität hat Calcium noch weitere Funktionen im Zellstoffwechsel- Erregung von Nerven- und Muskelzellen, Beteiligung am Glykogen-Stoffwechsel und der Zellteilung sowie Bestandteil bei der Aktivierung von Enzymen und Hormonen. Die empfohlene Tagesdosis liegt bei Erwachsenen bei 1000 mg. Mit den Acai Beeren gibt es eine natürliche Calciumquelle. Für eine gute Aufnahme des Calciums in den menschlichen Organismus ist eine ausreichende Versorgung mit Vitamin D3 notwendig.
- Über 40% der Beere bestehen aus Ballaststoffen: Diese unterstützen die Verdauung und kurbeln den Stoffwechsel an. Eine gesunde Verdauung stärkt zudem das Immunsystem. Durch die Unverdaulichkeit von Ballaststoffen wird das Nahrungsvolumen vergrößert, ohne jedoch den Energiegehalt zu steigern. Es tritt eine schnelle Sättigung ein. Dies wird dazu genutzt, die Acai Beere als Diätmittel zu bewerben und ihr eine Zügelung des Appetits nachzusagen. Jedoch gibt es keine Nachweise dazu, in welchem Maße sich dieser Effekt auf den Organismus auswirkt.
- Vitamin B: Die Acai Beere beinhaltet Vitamin B1 (Thiamin), B2 (Riboflavin), B3 (Niacin) und B6(Pyridoxin). Dabei handelt es sich um wasserlösliche Vitamine, die im menschlichen Organismus als Coenzyme und prosthetische Gruppen in Enzymen wirken. Vitamin B1 ist wesentlicher Bestandteil im Kohlenhydrat-Stoffwechsel und wichtig für Schilddrüse sowie Nervenfunktionen. Vitamin B2 entfaltet seine Wirkung bei der Konzentrationsfähigkeit und ist gegen Migräne. Bei der Verwertung von Fetten, Eiweißen und Kohlenhydraten sowie für schöne Haut und Nägel wird Vitamin B3 benötigt. Einen Schutz vor Nervenschädigungen leistet Vitamin B6.
- Vitamin C: wirkt ebenfalls als Radikalfänger und dient als Schutz vor Infektionen. Zusammen mit Vitamin B3 und B6 wird die L-Carnitin-Produktion gesteuert, welches für die Fettverbrennung in den Muskeln benötigt wird.
- Vitamin D: ist wichtig für die Calciumaufnahme in den Körper.
- Vitamin E: wirkt als Radikalfänger und ist für die Hemmung entzündlicher Prozesse, zur Stärkung des Immunsystems und für die Zellerneuerung.
- Omega-3- und Omega-9- Fettsäuren: Diese Fettsäuren schützen das Herz und haben einen positiven Einfluss auf das gesamte, kardiovaskuläre System.
Acai Beeren enthalten weiterhin eine Vielzahl von Mineralien, wie Magnesium, Zink, Kupfer und Kalium. Mineralien sind im Körper zum einen am Aufbau als Mengenelemente beteiligt und erfüllen zum anderen wichtige Funktionen als Spurenelement. Magnesium, Zink und Kupfer sind wichtige Bestandteile von Enzymen. Magnesium und Kalium haben zudem wichtige Aufgaben in der Erregungsweiterleitung.
Polyphenole wie Catechin und Epicatechin, Quercetin und andere Flavonoide: Wie andere Antioxidantien wirken auch diese Polyphenole entzündungshemmend und können Krebs vorbeugen. Eine Fettablagerung in den Blutgefäßen kann vermindert und einer Arteriosklerose kann vorgebeugt werden.
Der gesamte Mix an verschiedenen Inhaltsstoffen macht die Acai Beere bedeutend für den menschlichen Organismus. Die einzelnen Stoffe verhalten sich synergistisch im Stoffkreislauf, d.h. sie beeinflussen sich gegenseitig bzw. wirken miteinander.
Acai-Beere kaufen
Acai-Beere zu kaufen ist gar nicht so einfach. Zwar ist sie als Trendfood in verarbeiteter Form praktisch überall verfügbar. Jedoch ist nicht jedes Acai-Beere-Produkt tatsächlich sein Geld wert.
Einige Tests der Verbraucherzentralen haben ergeben, dass mit der Acai-Beere besonders viel Schindluder getrieben wird. Pestizidrückstände übertreffen in den getrockneten und gepulverten Beeren regelmäßig jeden Grenzwert. Bei billigen Produkten wurden auch schon mal Schimmelsporen nachgewiesen. Andere Importeure lügen dreist mit falschen Prüflaboren oder bei den Herkunftsländern. Viele Händler versuchen, den Hinweis „Made in China“ zu umgehen und geben fälschlicherweise andere Herkunftsländer, wie „Thailand“ oder „Tibet“ an.
Beim Acai-Beeren Kauf besteht deshalb ein besonderer Bedarf an Obacht. Das Schnäppchen von der Online-Handelsplattform ist nicht umsonst so günstig. Vertrauenswürdige Quellen für Acai-Beere sind bislang noch am Ehesten die Apotheken, Drogerien, Reformhäuser oder Onlines-Shops, die sich schon längere Zeit am Markt behaupten konnten.
Bedauerlicherweise gibt es die Acai-Beere hier in Europa praktisch nur in verarbeiteter Form: Getrocknet und zu Pulver zermahlen hat die Beere schon einen Teil ihrer wertvollen Inhaltsstoffe verloren. Es bleibt zu hoffen, dass es auch den Züchtern hierzulande gelingt, die Beere anzubauen. Das würde viele Probleme rund um die Acai lösen.
[amazon box=“B0767D3Y26″]
Fazit
In Studien konnte nicht nachgewiesen werden, dass die Acai Beere das Schlankheitsmittel ist, für welches es beworben wird. Es sind jedoch viele Inhaltsstoffe der Beere entschlüsselt worden, die in synergistischer Weise positive Auswirkungen auf den menschlichen Organismus und seinen Stoffwechsel haben. Besonders hervorzuheben sind die Antioxidantien, die durch das Abfangen der freien Radikale die menschlichen Zellen schützen. Es findet eine gesamte Stärkung des Körpers statt.
Das könnte Sie auch interessieren

Von der Natur inspiriert: Wie Eurycoma longifolia Deine Fitness und Vitalität steigern kann
Von der Natur inspiriert: Wie Eurycoma longifolia Deine Fitness und Vitalität steigern kann Bist Du auf der Suche nach natürlichen Wegen, um deine Fitness und Vitalität zu steigern? Dann könnte Eurycoma longifolia die Lösung sein, nach der du gesucht hast. Dieses...

Olivenöl: Wertvolle Inhaltsstoffe für einen abwechslungsreichen Essensplan
Wer an eine gesunde Ernährung denkt, denkt vielleicht nicht unweigerlich an Öl. Hochwertiges Olivenöl ist jedoch ein guter Beweis dafür, dass es eine gute Idee sein kann, hier ein wenig über den Tellerrand hinaus zu denken – und unter anderem vielleicht auch neue...

Die natürliche Heilung der Thai Massage
Was ist eine Thai Massage? Die aus Indien stammende Thai Massage existiert bereits seit über 2.500 Jahren. Ursprünglich als Heilkunst angesehen, beinhaltet die traditionelle Thai Massage Einflüsse sowohl aus der ayurvedischen als auch aus der traditionellen...
Bienenwachs zur Herstellung von Duftkerzen
Bienenwachs zur Herstellung von DuftkerzenKerzen selbst herstellen ist gerade im Herbst sehr beliebt. Kalte und regnerische Tage kann man dabei nutzen, um mit den Kindern außergewöhnliche Kerzen herzustellen und diese vielleicht an Weihnachten zu verschenken. Doch...
Moor
Wissenswerte Informationen über Moore im Überblick Bei einem Moor handelt es sich um ein feuchtes Gebiet, welches sich meistens über einige Meter bis Kilometer erstreckt. Besonders zeichnen sich diese Gebiete durch schwammige Böden aus, die nicht austrocknen. Aufgrund...
Vitalität und Gesundheit mit natürlichen Mitteln
Vitalität und Gesundheit mit natürlichen MittelnWir alle kennen das Phänomen, wenn wir zum Arzt gehen, um uns durchchecken zu lassen. Gesund geht man in die Praxis und mit zahlreichen Krankheiten kommt man wieder heraus. Damit dies aber erst gar nicht der Fall ist,...
Detox Kur
Detox Kur und was steckt dahinter? | Anleitung, PlanImmer öfter hört man mittlerweile von der Detox Kur und dass zahlreiche Blogger Selbstversuche unternehmen, um sie dann mit ihren Lesern zu teilen. Aber was ist das überhaupt und was macht man dabei? Ist es sinnvoll...
Lachsöl
Lachsöl für Mensch und TierLachsöl ist gesund und ein Jungbrunnen für den ganzen Körper. Doch nicht nur Menschen profitieren von dem hervorragenden Öl, sondern auch auf die Haustiere hat das Lachsöl eine positive Wirkung. Lachsöl ist voller gesunder Fettsäuren und ist...
Weitere nützliche Links
Wer schreibt hier
Natur-Institut Autor
Unser Autor R. S. ist Diplom Medienwirt (M.A.) und hat an der Universität Siegen studiert. Durch Zusatzqualifikationen zum Maschinenbautechniker und technischen Redakteur konnte er sich ein umfassendes Wissen rund um Maschinen, Anlagen und Sensoren aneignen. Sein Hauptberuf ist das Verfassen von Betriebsanleitungen, Arbeitsbeschreibungen, Produktdatenblätter und Informationsbroschüren. Die dort notwendigen Qualifikationen der genauen Recherche und der verständlichen Formulierung setzt er auch gewinnbringend bei seiner Autorentätigkeit ein.
Die freiberufliche Autorentätigkeit übt er seit 2013 aus. Die genaue Recherche von Informationen, das Aufbereiten von Sachverhalten und die Formulierung zu gut lesbaren Texten ist ihm eine Leidenschaft. Zum Thema „Manuka Honig“ ist Herr S. über seine Freiberuflichkeit gestoßen und konnte bereits hunderte gut recherchierter und exzellent formulierter Texte zu diesem Thema abliefern. Glasklare, unmissverständliche Informationen, gewürzt mit interessanten Hintergrund-Informationen, pfiffigen Tipps und einer kleinen Prise Wortwitz sind das Rezept, welche die Texte von Herrn S. so erfolgreich machen. Wir arbeiten seit zwei Jahren mit Herrn S. zusammen und freuen uns nach wie vor jedes Mal auf´s Neue auf seine Beiträge.
Quellenangabe
Catherine Ulbricht, Ashley Brigham u. a.: An Evidence-Based Systematic Review of Acai (Euterpe oleracea) by the Natural Standard Research Collaboration] In: Journal of Dietary Supplements.9(2), 2012, S. 128–47, doi:10.3109/19390211.2012.686347.
S. R. S. Cardoso, N. B. Eloy u. a.: Genetic differentiation of Euterpe edulis Mart. populations estimated by AFLP analysis (PDF; 565 kB), In: Molecular Ecology. 9, 2000, S. 1753–1760, PMID 11091311.
Barbosa RL, Dias VL, Pereira KS, Schmidt FL, Franco RM, Guaraldo AM, Alves DP, Passos LA (2012): Survival In Vitro and Virulence of Trypanosoma cruzi in Açaí Pulp in Experimental Acute Chagas Disease. Journal of Food Protection, 75 (3), S. 601–606.
Consumers Warned of Web-Based Açai Scams Center for Science in the Public Interest vom 23. März 2009, abgerufen 29. November 2009.
Brazilian Acai Berry Antioxidants Absorbed By Human Body auf sciencedaily.com vom 17. Oktober 2008, abgerufen 29. November 2009.
What are acai berries, and what are their possible health benefits? auf Mayo Clinic vom 23. Mai 2008, abgerufen 29. November 2009.